


High-throughput single-cell sequencing is mostly based on the use of cell barcodes. However, in certain cases, it can be superfluous and impractical, as every cell needs to be presented with a unique set of DNA oligonucleotides. This is achieved by single-cell compartmentalization into barcode-bearing droplets [1-3], nanowells[4], or by split-and-pool barcoding in regular multi-well plates [5]. The obtained sequencing data is interpreted using the ‘1 barcode = 1 cell’ paradigm: all reads bearing the same barcode are considered to originate from the same cell.
Barcoding-Free Multiplex Single-Cell Amplicon Sequencing
Rather than being linked by barcode information, DNA targets originating from the same cell can be concatenated into a single DNA fragment, leading to a ‘1 read = 1 cell’ data interpretation: all information within a single physical read is considered to be originating from the same cell. A notable example of the ‘1 read = 1 cell’ principle is emulsion overlap PCR to produce a single fusion amplicon from two single-cell-derived DNA targets. This approach has shown efficacy in diverse applications including chromosomal haplotyping [6], T-cell receptor chain pairing [7], and the association of phylogenetic markers with functional genes in bacteria [8]. Notably, EpicPCR [8] and OIL-PCR [9] have been employed to link 16S rRNA variants to information contained in extrachromosomal plasmids. This capability is crucial for facilitating high-throughput investigations into the propagation of antibiotic resistance genes via horizontal gene transfer.
Despite its potential, emulsion extension PCR is not scalable beyond two targets per cell, and working with droplets is challenging due to the difficulty in combining multiple molecular biology steps necessary to produce a fused PCR amplicon. This is particularly pronounced when working with microbes that require harsh lysis conditions.
Our semi-permeable capsule technology benefits from the throughput of droplets, enabling the study of thousands to hundreds of thousands of single cells per experiment. In contrast to droplets, SPCs enable full reagent exchange between consecutive reactions without loss of nucleic acid compartmentalization.
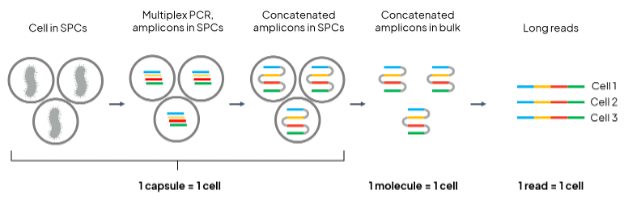
Figure 1. Schematic depiction of single bacteria isolation and read concatenation principle.
To demonstrate that SPCs allow extending the ‘1 read = 1 cell’ principle beyond two microbial targets (Fig. 1), we used a mixture of E. coli (transformed with the pUC-GFP vector) and B. subtilis cells encapsulated in SPCs using the Onyx microfluidic system. One chromosomal (16S rRNA-encoding) and two extrachromosomal (GFP-encoding and AmpR-encoding) genes were amplified from individual cells and concatenated within the SPCs. Next, SPCs containing approximately 8,000 single-cells were dissolved, releasing their contents, from which two Nanopore sequencing libraries were prepared: one for 16S-only sequencing, and the other enriching for the full GFP-AmpR-16S concatemer. In the 16S-only library, approximately equal numbers of reads mapping to E. coli 16S and B. subtilis 16S are expected. In the GFP-AmpR-16S library, only E. coli 16S should be observed.
The expected results were experimentally confirmed: 1) in the 16S-only library, 53% of length-filtered reads mapped to E. coli 16S rRNA, and 47% to B. subtilis 16S rRNA; 2) in the GFP-AmpR-16S library, 92% of length-filtered reads mapped to E. coli 16S rRNA.
The compatibility of SPCs with multi-step molecular biology workflows is well exemplified in this proof-of-concept, which included harsh alkaline lysis, PCR, proteinase K and USER enzyme mix treatment, and ligation, with straightforward washes between the individual steps. Notably, all steps were carried out on thousands of cells in parallel in a single 1.5-ml tube.
References:
- 1. Macosko EZ, Basu A, Satija R, Nemesh J, Shekhar K, Goldman M, Tirosh I, Bialas AR, Kamitaki N, Martersteck EM, Trombetta JJ, Weitz DA, Sanes JR, Shalek AK, Regev A, McCarroll SA. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell. 2015;161(5):1202-1214.
- 2. Klein AM, Mazutis L, Akartuna I, Tallapragada N, Veres A, Li V, Peshkin L, Weitz DA, Kirschner MW. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell. 2015;161(5):1187-1201.
- 3. Clark IC, Fontanez KM, Meltzer RH, Xue Y, Hayford C, May-Zhang A, D’Amato C, Osman A, Zhang JQ, Hettige P, Ishibashi JSA, Delley CL, Weisgerber DW, Replogle JM, Jost M, Phong KT, Kennedy VE, Peretz CAC, Kim EA, Song S, Karlon W, Weissman JS, Smith CC, Gartner ZJ, Abate AR. Microfluidics-free single-cell genomics with templated emulsification. Nat Biotechnol. 2023;41(11):1557-1566.
- 4. Goldstein LD, Chen YJ, Dunne J, Mir A, Hubschle H, Guillory J, Yuan W, Zhang J, Stinson J, Jaiswal B, Pahuja KB, Mann I, Schaal T, Chan L, Anandakrishnan S, Lin CW, Espinoza P, Husain S, Shapiro H, Swaminathan K, Wei S, Srinivasan M, Seshagiri S, Modrusan Z. Massively parallel nanowell-based single-cell gene expression profiling. BMC Genomics. 2017;18(1):519.
- 5. Yin Y, Jiang Y, Lam KG, Berletch JB, Disteche CM, Noble WS, Steemers FJ, Camerini-Otero RD, Adey AC, Shendure J. High-Throughput Single-Cell Sequencing with Linear Amplification. Mol Cell. 2019;76(4):676-690.e10.
- 6. Turner DJ, Hurles ME. High-throughput haplotype determination over long distances by haplotype fusion PCR and ligation haplotyping. Nat Protoc. 2009;4(12):1771-83.
- 7. Turchaninova MA, Britanova OV, Bolotin DA, Shugay M, Putintseva EV, Staroverov DB, Sharonov G, Shcherbo D, Zvyagin IV, Mamedov IZ, Linnemann C, Schumacher TN, Chudakov DM. Pairing of T-cell receptor chains via emulsion PCR. Eur J Immunol. 2013;43(9):2507-15.
- 8. Spencer SJ, Tamminen MV, Preheim SP, Guo MT, Briggs AW, Brito IL, A Weitz D, Pitkänen LK, Vigneault F, Juhani Virta MP, Alm EJ. Massively parallel sequencing of single cells by epicPCR links functional genes with phylogenetic markers. ISME J. 2016;10(2):427-36.
- 9. Diebold PJ, New FN, Hovan M, Satlin MJ, Brito IL. Linking plasmid-based beta-lactamases to their bacterial hosts using single-cell fusion PCR. Elife. 2021;10:e66834.